Expression pattern and subcellular localization of the VvHB58 protein
Previous studies analyzed the seed transcriptomes of progeny derived from the seedless paternal parent ‘Centennial seedless’ (V. vinifera) and seeded maternal parent ‘Red Globe’ (V. vinifera) to identify genes related to seed development. Transcriptome data revealed some homeobox genes may be involved in seed development process, and VvHB58 gene (Accession number: GSVIVT01008065001; GenBank: CBI15277) showed a differential expression pattern in the seeds of seedless and seeded progeny [11]. In this study, we used another four grape cultivars (two 3-year-old seedless cultivars ‘Thompson Seedless’ and ‘Flame Seedless’, and two 3-year-old seeded cultivars ‘Red Globe’ and ‘Kyoho’) to verify the expression pattern of VvHB58, so as to explore whether this differential expression pattern is universal. Previous studies have reported that the mass of the seeds in seedless grapes generally begins to decrease at 27 - 33 days after full bloom (DAF) [11]. We used qPCR to analyze the expression levels of VvHB58 during seed development at 27, 30, 33, 36, 39 and 42 DAF in these four cultivars (Fig. 1a). VvHB58 showed higher expression in the two seedless cultivars relative to the seeded cultivars. In order to further explore the expression pattern of VvHB58 in seed development stages, we selected 12 seed development stages to evaluate the expression of VvHB58 in 4-year-old ‘Thompson Seedless’ and ‘Red Globe’ (Because ‘Thompson Seedless’ and ‘Red Globe’ are representative cultivars of seedless grapes and seeded grapes respectively, the two cultivars were further considered for expression analysis in the following year). During seed development between 21 to 48 DAF, expression of VvHB58 was high in ‘Thompson Seedless’, but relatively low in ‘Red Globe’ (Fig. 1b), suggesting that VvHB58 may participate in seed development of seedless grapes.
In addition, we analyzed the expression pattern of VvHB58 in floral organs (stamen, pistil, petal and sepal) (Fig. 1c) and in the whole flower, root, stem, leaf, tendril and fruit (Fig. 1d) in the seedless cultivar ‘Flame Seedless’ and the seeded cultivar ‘Muscat Hamburg’. In general, the expression levels of VvHB58 in both vegetative and reproductive organs were higher in ‘Muscat Hamburg’ than in ‘Flame Seedless’, especially in the stamens, petals, sepals, roots, stems and fruit.
Consistent with its presumed role as a transcription factor, VvHB58 contains a putative nuclear localization sequence (NLS) of 10 amino acids, located at the beginning of the homeodomain (amino acids 68-78) (http://nls-mapper.iab.keio.ac.jp/cgi-bin/NLS_Mapper_form.cgi). To determine if VvHB58 might be localized in the nucleus, we fused the VvHB58 coding sequence to Yellow Fluorescent Protein (YFP) and expressed this transiently in tobacco (Nicotiana tabacum). As shown in Fig. 1e, the VvHB58-YFP protein accumulated in the nucleus.
Evolutionary analysis of VvHB58 protein
We previously reported the genome-wide identification of the homeodomain transcription factor family in the grapevine genome [27, 28]. An analysis of phylogeny and conserved domains identified members of the HD-Zip I family, including VvHB58. In this study, we further explored the evolutionary relationship between VvHB58, other grapevine HD-Zip I proteins, and HD-Zip I proteins from tomato [30], Arabidopsis [31], rice [32], soybean [33] and maize [34] (Additional file 1: Table S1). These HD-Zip I proteins can be divided into seven subfamilies: α, β, γ, δ, ε, ξ and φ (Fig. 2a). Interestingly, most HD-Zip I proteins from monocots including rice and maize clustered together in the ξ subgroup, while HD-Zip I proteins from dicots including Arabidopsis, tomato, soybean and grapevine were generally clustered together. The protein most homologous to VvHB58 in Arabidopsis is ATHB1, which has been reported to be involved in leaf development [35]. A homologous protein in tomato, LeHB1, has been reported to participate in fruit ripening and floral organogenesis [23]. Sequence alignment revealed that the homeodomain of VvHB58 shares 82.5% similarity with those of ATHB1 and LeHB1 (also known as SLHZ12) (Fig. 2b). The complete coding sequence of VvHB58 was amplified from seeds cDNA from the seedless grape ‘Thompson Seedless’ and the seeded grape ‘Red Globe’. This confirmed that genes from two cultivars encode identical proteins of 263 amino acids with a highly conserved homeodomain (amino acids 71 - 124) and a leucine zipper (amino acids 126 - 167).
Heterologous expression of VvHB58 in tomato causes a decrease in fruit size
To further investigate a potential role for VvHB58 in seed development, we expressed the VvHB58 gene in transgenic tomato (Fig. 3). Compared with nontransgenic tomatoes and a transgenic control only containing the empty vector, VvHB58 transgenic tomatoes were obviously dwarfed and exhibited reduced apical dominance (Fig. 3a). At the fruit ripening stage, transgenic VvHB58 lines showed smaller fruits with decreased seed number, but larger seeds (Fig. 3b-g). The number of VvHB58 transgenic tomatoes seeds was about half of that of non-transgenic tomatoes (Fig. 3f), but the weight of the transgenic tomatoes seeds increased slightly (Fig. 3g). It is speculated that VvHB58 may play a role in seed development. Although the number and weight of transgenic tomatoes seeds changed, the seeds developed normally. Additionally, we further analysed the fruit diameters and weights of transgenic and nontransgenic fruits during the fruit development and ripening process (Fig. 3c-e). During 10 - 70 DAF (from fruit set development to fruit ripening), both fruit diameters and weights of transgenic fruits were significantly smaller than nontransgenic type, suggesting VvHB58 gene may affect the early development of fruit sets, leading to a reduction of final cell number in fruits. Between 20 - 35 DAF, the weight of fruit from nontransgenic tomatoes increased rapidly, but that of fruit from VvHB58 transgenic lines increased only slowly.
Additionally, we analyzed the expression levels of VvHB58 and LeHB1 (the tomato homolog of VvHB58) in transgenic and non-transgenic tomatoes. VvHB58 was expressed especially strongly in seeds of the transgenic tomatoes. This is similar to the strong expression of VvHB58 seen during the seed development of seedless grapes (Fig. 3h). The transcript levels of LeHB1 gene involved in floral organogenesis and fruit ripening were not generally increased in the transgenic tomatoes (Fig. 3i), and LeHB1 gene only showed a higher transcript level in green fruits of transgenic tomatoes, suggesting ectopic expression of VvHB58 gene in transgenic tomatoes didn't result in increased expression of it's homologous genes in tomatoes.
Subsequently, we studied the floral organs and pollen grains of transgenic tomatoes. As evaluated by FDA staining, pollen viability was not significantly affected by VvHB58 expression (Fig. 4a). Additionally, we observed no difference in the size and shape of their pollen grains and floral organs (Fig. 4a). This suggests that VvHB58 gene regulates fruit and seed development may not via affecting stamens or pistils. Furthermore, we observed that the pericarp of the fruit from VvHB58 transgenic tomatoes was relatively thick compared with the control fruit. The pericarp cells of transgenic fruit (Fig. 4b) were smaller than controls, but the number of layers with small cells below the epidermis was increased, suggesting an additional role for VvHB58 in fruit enlargement prior to ripening. Histological longitudinal section of seeds of transgenic and control plants showed that the endosperm cells in transgenic seeds were larger and more loosely arranged, suggesting that the increased seed size may be at least partly due to larger endosperm cells (Fig. 4c).
Variations in phytohormone levels and hormone-related gene expression between VvHB58 transgenic and control plants
Seed development process in tomato is strongly tied to endogenous concentrations of auxin and gibberellin. High levels of these two hormones in the ovary play an important role in fruit set and early development [36]. To explore whether the observed effect of VvHB58 on the transgenic tomato fruit and seed is due to its influence on hormone signaling pathways, we measured the endogenous levels of the five classic phytohormones auxin, gibberellin, cytokinin, abscisic acid and ethylene in stems, leaves, flowers, pericarp, seeds, green fruit and ripening fruit, in both control and transgenic lines (Fig. 5a). We found that, in general, the content of endogenous auxin, gibberellin and ethylene were significantly different between transgenic and control plants. In contrast, cytokinin and abscisic acid levels showed no significant difference.
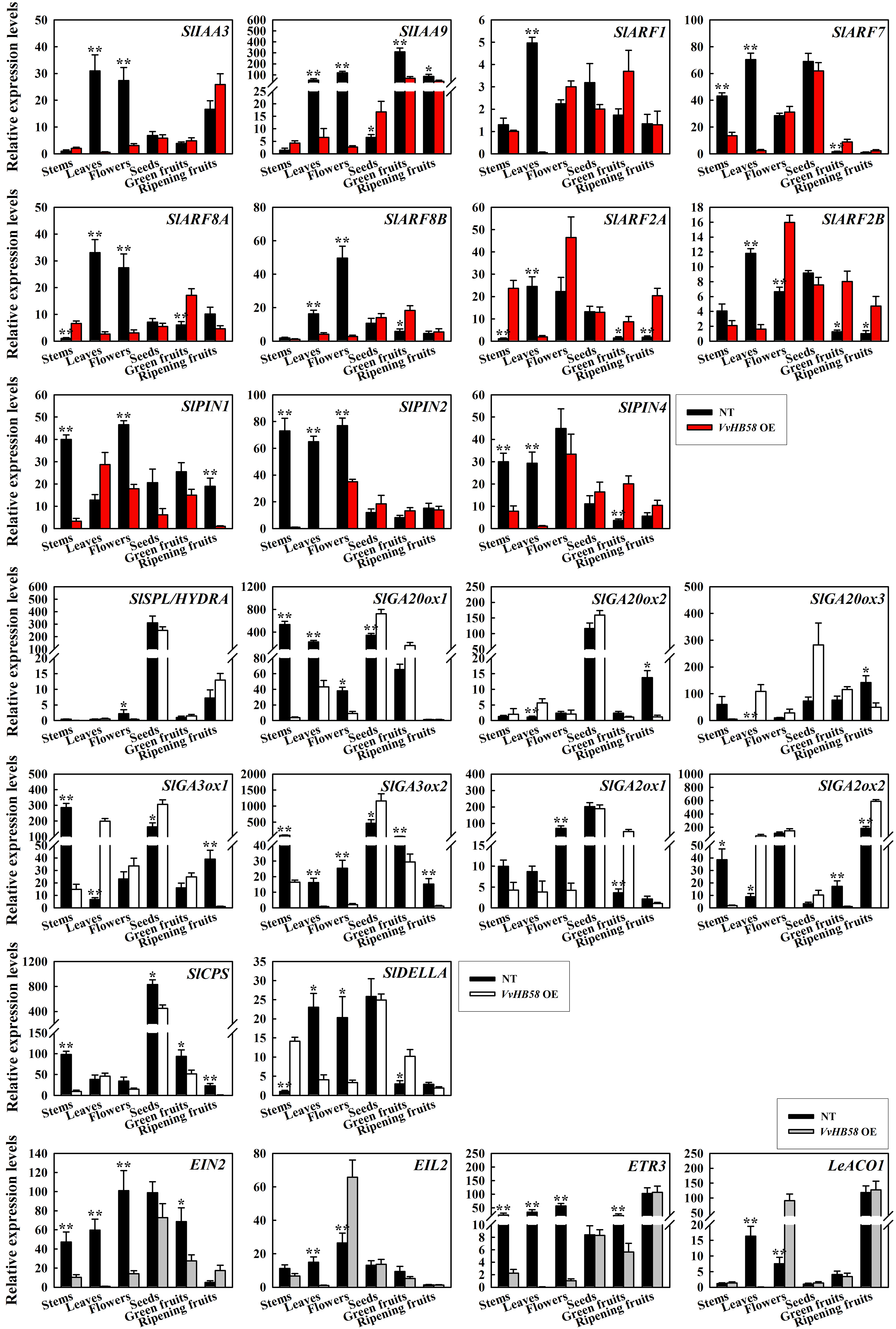
Except for flowers and ripening fruit, the content of endogenous auxin in transgenic tomatoes was significantly lower than that of control plants, and this is consistent with the the dwarfed stature and smaller fruit size of the transgenic tomatoes. Additionally, expression levels of several auxin responsive genes (SlIAA3, SlIAA9, SlARF1, SlARF7, SlARF8A, SlARF8B, SlARF2A and SlARF2B) described to be involved in the control of fruit initiation [36-38], and auxin distribution genes (SlPIN1, SlPIN2 and SlPIN4) in the SlPIN gene family known to be involved in fruit development were evaluated via qPCR [39, 40] (Fig. 5b and Additional file 2: Figure S1). In general, these genes showed stronger expression in control plants than in transgenic tomatoes. For example, the expression SlIAA3, SlARF8A and SlARF8B were higher in leaves and flowers of nontransgenic control plants compared with transgenic tomatoes. Likewise, SlPIN2 and SlPIN4, encoding PIN-like auxin efflux carriers, were expressed more strongly in stems, leaves and flowers of control plants. However, the expression levels of some genes did not coincide with the difference seen for auxin content. For instance, SlARF2A and SlARF2B showed increased expression in flowers and green fruit of transgenic tomatoes. This indicates that the regulatory effect of the VvHB58 gene on auxin may be carried out through a relatively complex network.
The content of endogenous gibberellin in stems, leaves, green fruit and ripening fruit of transgenic tomatoes was significantly lower than that of control plants, but was markedly higher in flowers, pericarps and seeds (Fig. 5a). This is consistent with the reduced gibberellin content in stems, leaves, green fruits and ripening fruit conditioning the dwarfing and smaller fruit size in transgenic tomatoes, and increased gibberellin content in flowers conditioning the observed decrease of seed number per fruit. Furthermore, the increase of gibberellin content in seeds of transgenic tomatoes may be connected with the larger seed size. The expression levels of genes involved in gibberellin biosynthesis, such as SlGA20ox1, SlGA20ox2, SlGA20ox3, SlGA3ox1, SlGA3ox2, SlCPS (copalyl diphosphate synthase) and SlSPL/HYDRA (SPOROCYTELESS/HYDRA) [36, 41], gibberellin inactivation SlGA2ox1 and SlGA2ox2 [42], and gibberellin response SlDELLA were also detected [43] (Fig. 5b and Additional file 2: Figure S1). Most of these genes showed stronger expression in seeds, and the five genes related to gibberellin biosynthesis showed higher transcript levels in seeds of transgenic tomatoes compared with control plants. In transgenic tomatoes, up-regulation of the gibberellin inactivation genes SlGA2ox1 and SlGA2ox2 in green and ripening fruit would be expected to reduce levels of bioactive gibberellin. In a word, the increased transcript levels of gibberellin biosynthesis genes and decreased transcript levels of gibberellin inactivation genes should increase bioactive gibberellin, which could promote premature ovary growth.
Tomatoes are climacteric fruit, and endogenous ethylene has a strong promotive effect on fruit ripening. In the VvHB58 transgenic tomatoes, the content of endogenous ethylene in stems, leaves, flowers, pericarps and seeds was dramatically lower than in control plants (Fig. 5a). We analyzed the transcript levels for the ethylene biosynthesis genes LeACO1 (1-aminocyclopropane-1-carboxylate oxidase 1), and ethylene receptor genes EIN2 (ethylene signaling protein), EIL2 (EIN3-like proteins) and ETR3 (NR, never-ripe) by qPCR [37] (Fig. 5b and Additional file 2: Figure S1). EIN2 and ETR3 exhibited similar transcript levels, showing lower expression in stems, leaves, flowers and green fruits of transgenic tomatoes compared with control plants. In contrast, EIL2 and LeACO1 showed relatively higher transcript levels in flowers of transgenic tomatoes. Altogether, considering both ethylene content changes and gene expression patterns, we concluded that ethylene responses are slightly impaired in the transgenic tomatoes.
Fruit and seed development is a complex yet tightly regulated process. Except for auxin, gibberellin and ethylene, it is also regulated by abscisic acid and cytokinin. The increased endogenous cytokinin content in transgenic seeds may lead to seed enlargement (Fig. 5a). More abscisic acid in the stems and leaves of transgenic tomatoes may promote premature senescence (Fig. 5a). On the whole, the regulatory effect of VvHB58 gene on fruits and seeds may be carried out mainly through changing endogenous auxin, gibberellin and ethylene content.
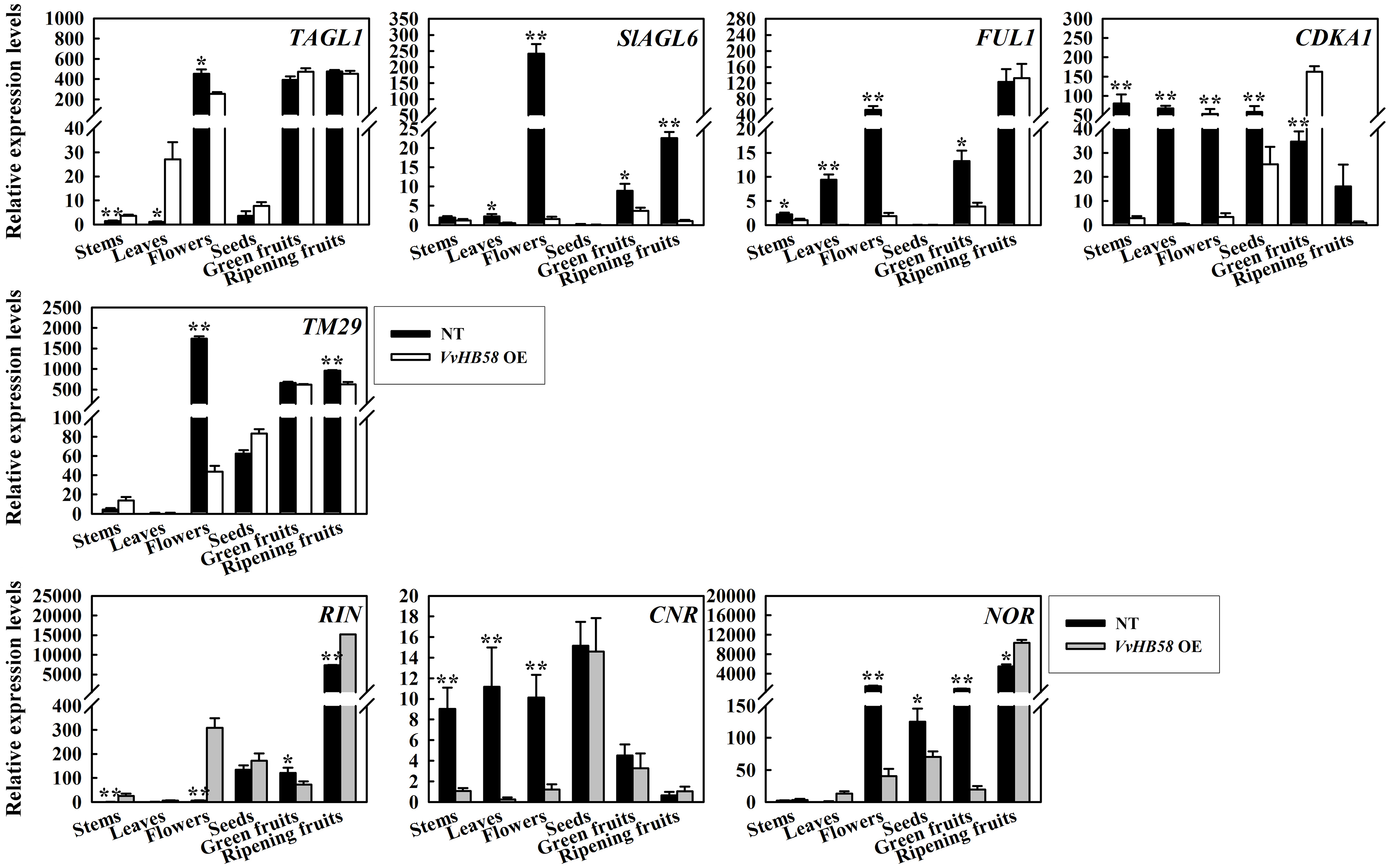
Additionally, we analyzed the expression levels of the fruit ripening-associated genes RIN (ripening inhibitor), CNR (colorless non-ripening) and NOR (nonripening) [37], and seed development-related gene TAGL1 (AGAMOUS-LIKE1), SlAGL6 (SlAGAMOUS-LIKE 6), FUL1 (FRUITFULL, previously called TDR4), CDKA1 (cyclin-dependent kinases) and TM29 (Tomato MADS-box 29) [44-48] (Fig. 5b and Additional file 3: Figure S2). In transgenic tomatoes, expression of NOR and RIN was strongly reduced compared with controls plants in green fruit, but was enhanced in ripening fruits. This indicates that ectopic expression of VvHB58 may cause a slight change in the fruit ripening mechanism. It has been reported that down-regulation of two genes, TM29 and SlAGL6, can lead to parthenocarpy in tomato [45, 48]. Consistent with our results, the transcript levels of these two genes were dramatically lower in the flowers and fruits of transgenic tomatoes compared with control plants. Parthenocarpy in tomato can also be caused by up-regulation of the CDKA1 gene [47]. Up-regulation of CDKA1 during early fruit development, concomitant with rapid cell division, may also lead to parthenocarpy. We found that the CDKA1 gene also exhibited higher transcript levels in green fruits of transgenic tomatoes. Furthermore, the TAGL1 gene involved in carpel and stamen development was down-regulated in the flowers of transgenic tomatoes [49], and FUL1 involved in fruit ripening exhibited a lower transcript level in flowers and green fruits of transgenic tomatoes. Taken together, this data suggests that VvHB58 may affect seed and fruit development-related genes to co-regulate growth and development process of seed and fruit.
Deletion analysis of the VvHB58 promoter
In order to elucidate the molecular basis of VvHB58 differential expression, we cloned the VvHB58 promoter from seeds DNA of ‘Thompson Seedless’ and ‘Red Globe’, including 1605 bp upstream from the ATG. We found that this sequence was invariant between the two cultivars. Analysis of this region for cis-acting elements identified several elements implicated in hormone responses, defense and stress responses, light responses and endosperm expression (Fig. 6a). Common among these were the GCN4 motif and Skn-1 motif related to endosperm expression, TGACG motif and CGTCA motif involved in methyl jasmonate responses, TCA element involved in salicylic acid responses, ABRE element associated with abscisic acid, HSE element involved in heat stress, MBS element related to drought inducibility, TC-rich repeats associated with defense and stress responses, LAMP element and AE-box involved in light response, and 5’ UTR Py-rich stretch conferring high transcription levels [27].
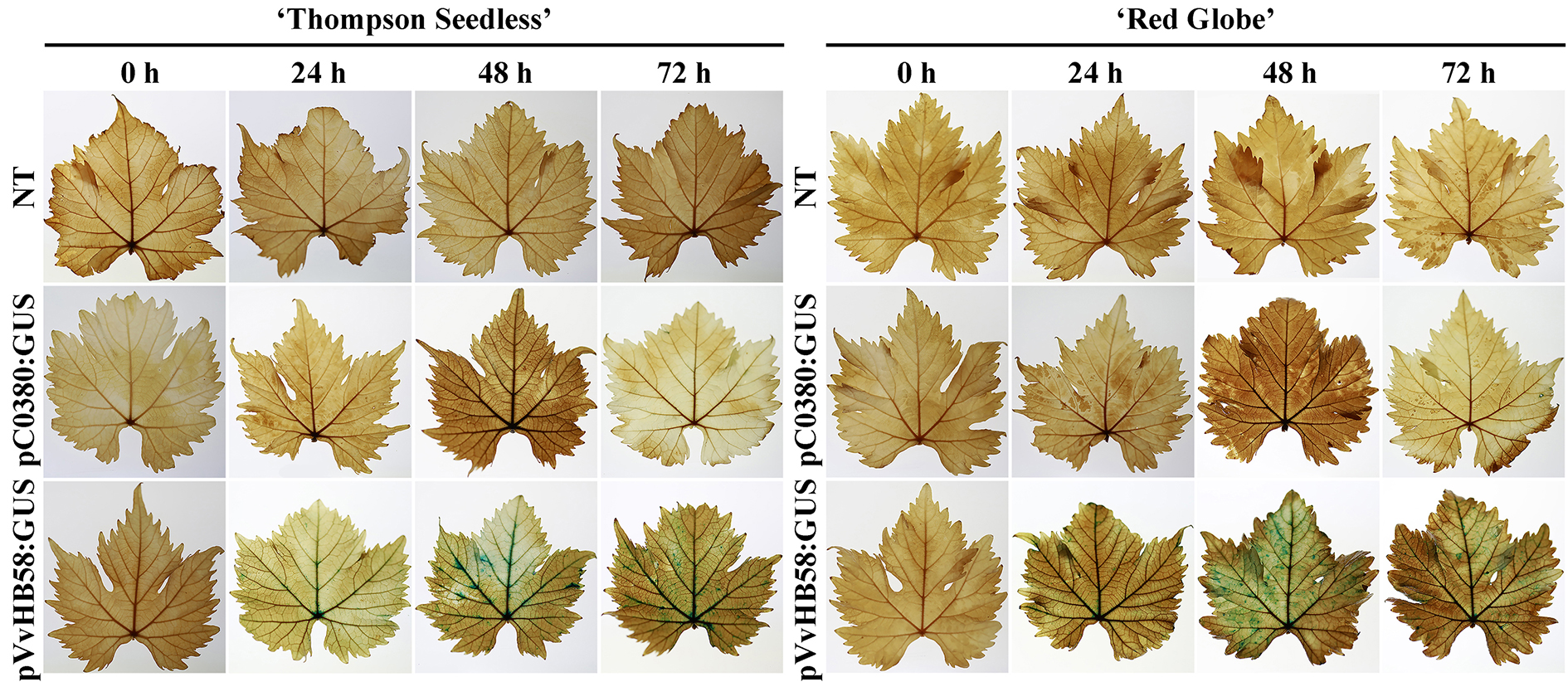
To identify which cis-elements are more important for VvHB58 transcription and if there are differences in VvHB58 promoter activity between seedless and seeded grapes, we carried out a histochemical assay of GUS (β-glucuronidase) expression in transiently transformed leaves of ‘Thompson Seedless’ and ‘Red Globe’. The 1605 bp VvHB58 promoter fragment was fused to a reporter gene encoding GUS, generating the construct pVvHB58:GUS, and a series of deletion fragments D1 (-1311 bp - ATG), D2 (-891 bp - ATG), D3 (-525 bp - ATG), D4 (-330 bp - ATG) were engineered. A construction with no promoter served as a negative control (pC0380:GUS) [50]. pVvHB58:GUS and pC0380:GUS were transformed into Agrobacterium tumefaciens and then introduced into grapevine leaves. The histochemical assay showed the GUS activity of pVvHB58:GUS reached the highest intensity at 48 h after A. tumefaciens infection (Additional file 4: Figure S3). Therefore, the subsequent assay used grapevine leaves at 48 h after infection. Additionally, no GUS activity was detected in nontransgenic control plants, and extremely little was detected for pC0380:GUS. All the deletion constructs were transformed transiently into leaves of ‘Thompson Seedless’ and ‘Red Globe’. Histochemical staining (Fig. 6b) and fluorometric analysis (Fig. 6c) of GUS activity indicated the highest level of GUS activity was detected in leaves of ‘Red Globe’ carrying the construction that included the entire 1605 bp VvHB58 promoter sequence. In contrast, GUS activity of the other four constructs (D2:GUS, D3:GUS and D4:GUS) showed higher intensity in ‘Thompson Seedless’ than ‘Red Globe’. Importantly, as deletion advanced between -1605 bp and -891 bp, GUS activity declined continuously in ‘Red Globe’, suggesting multiple cis-elements important for expression were being omitted. Nevertheless, the construct containing the deletion to the shortest fragment D4 (-330 bp - ATG) also showed a remarkable enhancement of GUS activity compared with D2 (-891 bp - ATG) and D3 (-525 bp - ATG) in both cultivars, indicating the shortest fragment D4 also has the ability to activate gene transcription. Together, these results suggest that both activating and repressive cis-elements are found in the VvHB58 promoter region.
Analysis of DNA methylation level in the VvHB58 gene
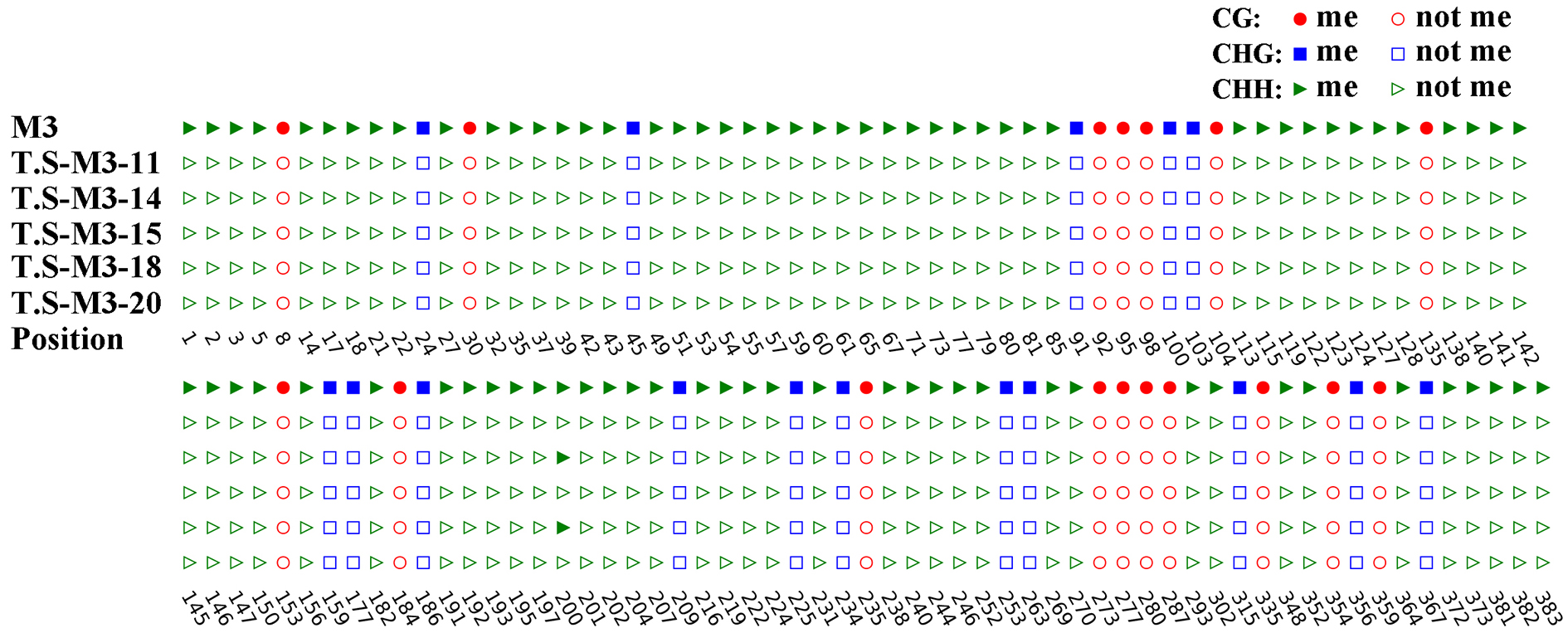
To investigate if DNA methylation might be involved in the differential expression of VvHB58 between seedless and seeded cultivars, bisulfite sequencing analysis was performed targeting three representative regions of the VvHB58 gene in ‘Red Globe’ (Fig. 7a). M1 (-1309 bp ~ -898 bp) and M2 (-955 bp ~ -499 bp) were located in the promoter, and M3 (-360 bp ~ 28 bp) was located in the promoter and first exon region. As shown in Fig. 7b, the DNA methylation level of the VvHB58 gene was very low, with only 40% CG type methylation in one CG context (-884 bp) of the M2 region. However, we found no cis-acting element near this CG site (Fig. 6a), suggesting that lower expression of VvHB58 in ‘Red Globe’ grape is independent of DNA methylation, and suggesting that other mechanisms may be responsible. However, it is also possible that regulatory changes in DNA methylation occurred outside of this identified region.
Expression patterns of genes encoding DNA methyltransferases and demethylases in seedless and seeded grape
Cytosine methyltransferases and demethylases play vital roles in maintenance of genomic methylation and are involved in various biological processes, such as gametogenesis [16, 18], apomixis [51] and fruit ripening [21]. To identify potential methyltransferases and demethylases in the grape genome scale, we carried out a homology-based search for sequences encoding grapevine cytosine methyltransferases and demethylases, using the NCBI-blastp (https://blast.ncbi.nlm.nih.gov/Blast.cgi) and using characterized methylation factors from Arabidopsis as queries. We identified and characterized the expression during seed development of methyltransferases VvCMT1 (XP_002275932.1), VvCMT2 (XP_019080798.1), VvCMT3 (XP_010651344.1), VvDDM1 (XP_002267239.2), VvDRM2 (XP_010660894.1), VvDRM3 (XP_019075700.1) and VvMET1 (XP_002267200.1), and demethylases VvDME (XP_002277401.1) and VvROS1 (CBI30244.3) (Fig. 8 and Additional file 5: Table S2).
In the period from 27-38 DAF the expression of VvCMT1, VvCMT3 and VvDDM1 were significantly higher in ‘Red Globe’ than in ‘Thompson Seedless’. Likewise, in the period from 34-38 DAF the expression levels of VvCMT2, VvDRM2, VvDRM3 and VvMET1 were higher in ‘Red Globe’ than ‘Thompson Seedless’. In contrast, the expression of VvROS1 and VvDME in ‘Thompson Seedless’ were higher than in ‘Red Globe’ during 27-32 DAF, then gradually declined during 34-38 DAF. In general, their expression levels gradually declined in ‘Thompson Seedless’ and gradually increased in ‘Red Globe’. The observation that grape seed development is related to changes in transcriptional levels of DNA methylation-related genes suggests that changes in DNA methylation may play a potential role in grape seed development.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}