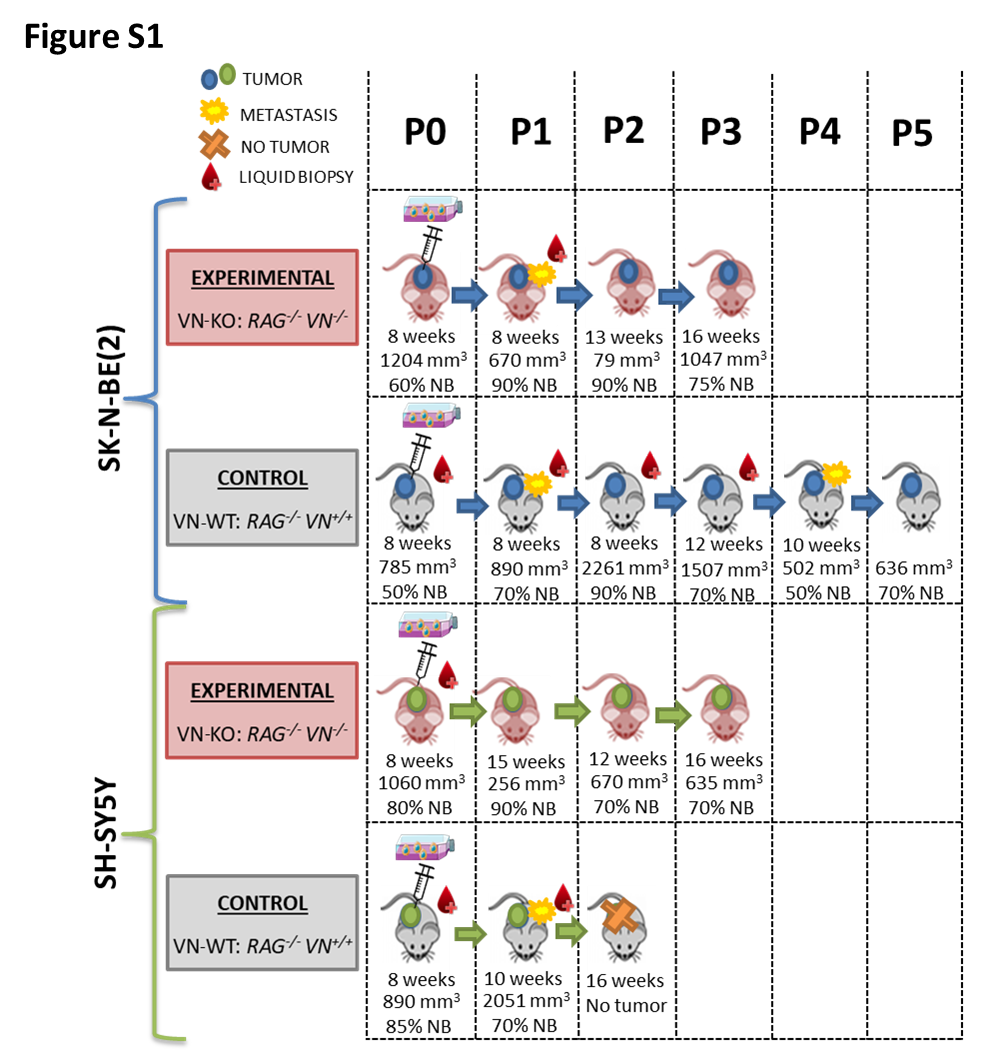
HR-NB is a disease with risky spatial-temporal genetic heterogeneity and high chromosome instability related to poor prognosis and therapy resistance [49, 50]. It is not yet clear whether this genomic variability could be the cause or consequence of selective evolutionary pressure in the tumor microenvironment[11]. We assumed that clonal genetic selection and/or evolution of aggressive NB cells could be promoted in part by the stiffness and composition of their surrounding matrix. To get closer to confirming this hypothesis, we selected two representative NB cell lines characterized by genomic alterations typically observed in HR-NB (MNA in SK-N-BE(2)[42] and ALK mutation in SH-SY5Y)[47]. Both cell lines were grown separately in hydrogels with ECM rigidity similar to the ECM of human HR-NB and xenograft tumors [39]. To make the 3D models even more biomimetic, malignant neuroblasts were cocultured with 10% Schwann cells in some cases, since even in poor stroma NB a low proportion can be found accompanying the undifferentiated or poorly differentiated neuroblasts [51]. To analyze the impact of ECM properties on genomic heterogeneity, we selected the HD-SNPa approach, complemented by the NGS technique, as one of the most commonly used procedures for detecting both MNA and SCAs in routine genetic diagnosis of NB [52]. Unlike this study, most research related to ECM tumor mechanotransduction has focused on uncovering changes in gene expression, which are difficult to translate to the clinic [53–55]. We looked for typical and/or atypical HR-NB SCA changes that could be associated with NB physics, and rapidly used for clinical translation to diagnosis and therapy. We also explored whether MNA and/or ALK-mutated cells respond similarly to ECM properties.
In previous studies, we demonstrated that elevated VN secretion by tumor cells was founded in rigid ECMs, and was related to poor patient outcomes and to growth of orthotopically inoculated NB cell lines[22]. Moreover, we observed that the high amount of VN is secreted forming tracks and we postulated that VN participates in stiffness, mediating biotensegrity and promoting tumor migration in HR-NB[22, 56]. We also reported that SK-N-BE(2) cells grown in stiff 3D hydrogels show more efficient adaptation, increased cell proliferation, high expression of the Bcl2 antiapoptosis marker, and high mRNA processing rate(24). In the present study, genomic analysis of SK-N-BE(2) cell line revealed a number of equivalent aberrations in VN-KO tumors and in cells grown in stiffest and/or longer cultured times hydrogels. In these samples, we observed a positive selection of cells that in addition to MNA, contained: i) stable SCAs (1p-, 3p- and 17pq-); ii) SRO generated from previous SCAs (+ 2p and chromotripsis-like of chromosome 21) and iii) new SCAs and FSCAs (1p-, +1q, 9p-, +9p, 9q-, +9q). We also observed negative pressure which triggered progressive loss of the remaining SCAs (+ 7q, + 11q, 13q-, 19q-, 20p-). Schwann cells seem to enhance clonal selection in NB in a more biomimetic context; however, the genomic differences found between hydrogels with vs without Schwann cells are minor and affect only the percentage of cells with some SCAs. The evolutionary adaptation we have detected may be due only to deterministic events (clonal selection) or also to interaction with stochastic processes (generation of mutations and genetic drift). However, the fact that genomes in two independent models, both with tumor spatial structures related to cellular aggressiveness, show similar genomic aberrations suggests a predominance of the deterministic evolutionary force. The increase in selection pressure, driven by adaptation to niches deliberately altered (by their lack of systemic VN, and by high stiffness and/or long 3D growth time) in both models, could prompt this similar clonal selection towards genetic aberrations detected by probably advantageous cellular phenotypes[57]. Both niches, which initially lacked VN, showed a large amount of VN secreted by neuroblats [22]. Our results show new evidence for the probably role of VN in biotensegrity mechanotransduction by mediating the cellular response to ECM stiffness [22, 56]. The adaptation process was faster in VN-KO tumor cells but less manageable than in hydrogels. Furthermore, cells grown in rigid 3D hydrogels showed faster selection for SCAs and less presence of intratumoral heterogeneity than those cells with long and soft 3D growth, which could reflect a more efficient adaptation to niche [39] as occurs in aggressive patients tumors with rigid ECMs[58]. Conversely, in 3D hydrogels with short culture times, cells may had not yet been subjected to the selective process, due to their incipient adaptation to ECM. As in the early stages of patient tumors, cells in 3D bioprinted hydrogels need to grow and survive, and then adjust genetically and epigenetically to acquire the features they need to take on new roles such as migration. For this among other characteristics, 3D models are increasingly important, reducing the use of animal models and controlling the parameters focused on the study, adding degrees of complexity to gain deeper insight into the tumor microenvironment.
Interestingly, some genes of the new SCAs undergoing positive selection in VN-KO tumors and stiff and/or long-cultured hydrogels are involved in ECM composition and architecture, mechanotransduction and cell migration. In chromosome 1 there is special focus on the collagen gene COL11A1 (in the nearer centrometic fragment of 1p-), the genes related to actin ARPC5 and ACTN2, some laminins and the integrin α10 (in + 1q), while the deleted FSCA of chromosome 9p contains the genes KANK1 and DOCK8, both involved in actin polymerization [59–61]. In our customized NB-mechanopanel, COL11A1 and DOCK8 genes specifically showed certain gene variants that have disappeared in the tumors from VN-KO mice with the mentioned heterozygote deletions. The extent to which this occurred, even when the cell population with these mutations was not inconsiderable in the other growing conditions (allelic frequency: 0.4), raises the question of whether these variants might play a role in adaptation. VN of ECM could have a swift impact on the mutational profile of these migration-related genes.
Chromosome 9 aberrations have not been a focus of interest so far in HR-NB, unlike those of chromosome 1 (1p- and + 1q) [32, 62, 63]. Therefore, we decided to delve on 9 chromosome abnormalities in a cohort of primary human HR-NB. Chromosome 9 aberrations involving KANK1 and/or DOCK8 genes were detected in 39.53% of the MNA primary HR-NB analyzed. Consequently, we recommend a spotlight on chromosome 9 aberrations[64–66] affecting DOCK8 and KANK1 in NB, and especially on the sometimes denigrated FSCAs, as they could involve invasion in MNA HR-NB, as occurs in other malignant tumors [67]. The relevance of these new genetic alterations found in human HR-NB, especially the frequency and prognostic impact of DOCK8 and KANK1, should be determined in collaborative studies.
The SK-N-BE(2) cell line has been characterized as particularly heterogeneous[42]. Likewise, high genetic instability of xenograft mice, showing gains and losses of SCAs with passages, has already been described[57]. In all our in vitro and in vivo samples only two SCAs remained stable (with the same length and break points) in most cells: partial deletion of 3p and 17pq, adding MNA. 3p loss is a common event in HR-NB[68]; the region contains certain important suppressor genes, including RHOA. RhoA protein plays an significant role in mechanotransduction, as it mediates focal adhesions and stress fiber formation[69]. Its reduced signal (associated with 3p) has been linked to growth and metastasis in colorectal tumors [70, 71]. Moreover, in these tumors RHOA downregulation is used to match the activation of Wnt signal pathway and inactivation of TGFβ. All together they participate in initiation and progression of tumors [70]. According to Gene Onthology, the Wnt pathway is one of the most affected by genomic aberrations of SK-N-BE(2) cells, and most of the altered genes of these pathways are in SCAs that changed (were positive or negative selected) in VN-KO tumors and 3D bioprinted models (+ 7q, +9q, + 11q, 13q-). TGFβ is located in the 19q deleted region which also had reduced presence in the samples mentioned. This gene has dual action: progression of tumors when underexpressed, and degradation of the ECM when its levels increase[72]. Meanwhile, 17pq- contains the tumor suppressor gene TP53, which plays an important role in tumorogenesis and aggressiveness[73]. This gene was also mutated in all samples from SK-N-BE(2) cell line with an alleleic frequency close to 1. The double mechanism involving a mutation in one of the alleles and a deletion or CNLOH of the other one has been highlighted for its combined negative impact [73]. In addition, the reported imbalance of chromosome 17, in which a loss of 17pq has been detected, is usually found with the gain of the remaining fragment of the chromosome, that is, a typical + 17q[74]. 17q gain is precisely the most common genomic aberration of NB [31]. In fact, the hidden + 17q became evident in some of our hydrogels and in VN-WT tumors. A probable explanation for this is that in addition to overexpression of tumor progression–promoting genes within the 17q fragments, the loss of function of genes localized on 17p- or CNLOH of 17p (such as TP53) may play a significant role in neuroblastoma development[74].
The impact of ECM properties on genomic heterogeneity of the SH-SY5Y cell line was not evident when using our models. These cells may be less sensitive, or would adapt to ECM stiffness/composition differently, without SCA changes or longer-term SCA changes. Two research groups have described morphological and gene expression changes in this cell line in stiff environments [53, 54]. It is possible that ALK-mutated cells respond to ECM changes through epigenetic alterations or genetic mutations undetected by the techniques we employed[53, 75–77].
{kind=link}